양봉 뚜렷한 바이러스 구성과 바이러스 다각화의 부족은 바이러스 스필오버가 서부 꿀벌과 일반 동부 땅벌 사이의 막다른 길임을 나타냅니…
페이지 정보

본문
뚜렷한 바이러스 구성과 바이러스 다각화의 부족은 바이러스 스필오버가 서부 꿀벌과 일반 동부 땅벌 사이의 막다른 길임을 나타냅니다.
병원체의 종간 전파는 병원체가 이전 에 해당 병원체에 감염되지 않은 숙주에게 전파될 때 발생하며 , 그 결과 중 하나는 스필오버 로 , 병원체가 새로운 유지 숙주를 확립 하는 것입니다. 바이러스 스필오버는 관리되는 서부 꿀벌( Apis mellifera ) 군집에서 바이러스 스필오버가 발생할 가능성이 있기 때문에 야생 곤충 수분 매개자에게 위협이 되는 것으로 널리 간주됩니다. A. mellifera 에서 땅벌( Bombus 속)으로 의 바이러스 스필오버에 많은 관심이 집중되었는데 , 많은 땅벌이 개체수 감소를 겪고 있으며 일부 종은 멸종 가능성이 높습니다 .4 , 5 스필오버는 다면적인 현상이며 다양한 시나리오를 나타내는 데 사용되는 용어입니다 . 그림 1 에서 정의한 대로 노출, 막다른 스필오버, 범위 확장을 동반한 스필오버 라는 용어를 사용합니다 . 간단히 말해서, 첫 번째이자 가장 흔한 시나리오는 노출입니다. 노출은 실제 숙주와 공통 환경을 공유함으로써 생물체 내부 또는 표면에 바이러스가 우연히 존재하는 것으로, 감염과는 무관합니다. 새로운 숙주에 노출된 바이러스가 최소한 감염을 유발할 수는 있지만 새로운 숙주 간에 충분히 전파되지 못할 경우, 바이러스가 새로운 숙주 집단에서 필연적으로 제거되는 막다른 스필오버 2 가 발생합니다 (그림 1 ).
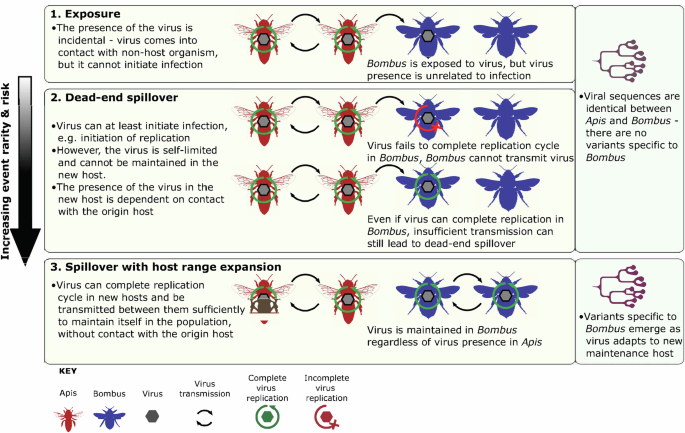
바이러스 스필오버에 관한 다른 프레임워크 1 , 2 , 3 , 6 , 7 , 8 을 기반으로 합니다 . Melissa Broussard가 제작한 Apis와 Bombus 실루엣은 https://www.phylopic.org/ 에서 가져왔으며, https://creativecommons.org/licenses/by/4.0 라이선스에 따라 허가를 받아 수정했습니다 .
새로운 숙주를 확립하기 위해 바이러스는 분자 상호작용, 숙주 생태 및 기타 생물학적 요인을 포함한 복잡한 장벽을 극복해야 합니다. 결과적으로, 노출 및 막다른 스필오버는 다른 동물 시스템에서 볼 수 있는 것처럼 범위 확장을 동반한 스필오버보다 훨씬 더 흔합니다 7 , 8 . 새로운 숙주를 확립하는 데 중요한 장벽을 극복할 수 있는 바이러스조차도 여전히 막다른 스필오버에 도달할 수 있습니다. 예를 들어, 새로운 숙주에서 복제를 완료하고 대량의 비리 온 을 생산할 수 있는 인플루엔자 바이러스조차도 새로운 유지 숙주를 확립하기에 충분한 바이러스 전파가 없을 수 있습니다 8 . 바이러스가 이러한 장벽을 극복하고 새로운 숙주 간의 전파를 통해 새로운 숙주 집단에서 자신을 유지할 수 있는 시나리오에 대해 범위 확장을 동반한 스필오버라는 용어를 사용합니다(그림 1 ). 바이러스는 자신을 유지할 수 있으므로 새로운 숙주에서 바이러스의 존재는 원래 숙주의 바이러스 저장소와의 접촉에 의존하지 않습니다.
범위 확장을 동반한 스필오버의 주요 특징은 병원체 9 의 진화가 높아지고 새로운 숙주에 적응한 변종이 빠르게 출현하는 것입니다(그림 1 ). 반면 노출 및 막다른 스필오버 시나리오에서는 바이러스의 전파 및 적응이 실패하여 바이러스 서열 동일성이 원래 숙주의 서열과 본질적으로 동일하게 유지됩니다. 꿀벌과 다양한 다른 절지동물 간의 스필오버를 제안하는 많은 연구가 있지만 10 스필오버 관련 시나리오는 잘 구분되지 않았습니다. 문헌에서 논의되는 우려 시나리오는 범위 확장 위협이 있는 스필오버인데, 새로운 유지 숙주의 확립이 숙주 개체군에 가장 큰 위협이 되기 때문입니다(그림 1 ). 그러나 대부분의 스필오버 증거는 바이러스 존재 평가에 기반을 두고 있으며, 노출, 막다른 스필오버, 범위 확장을 동반한 스필오버를 구분하는 능력이 제한적입니다. 결과적으로 노출이나 막다른 스필오버 사례가 범위 확장을 동반한 스필오버로 널리 잘못 특성화되었을 가능성이 높습니다. 꿀벌에서 발생하는 스필오버 관련 시나리오를 올바르게 식별하는 것은 수분매개자에게 다양한 정도의 위험을 초래하기 때문에 매우 중요합니다. 위험은 노출에서 막다른 스필오버, 분포 범위 확대에 따른 스필오버까지 증가합니다.
비교적 좁은 범위의 바이러스가 꿀벌 바이러스 스필오버의 맥락에서 연구되었으며, 주로 A. mellifera 바이러스 변형 날개 바이러스(DWV, Iflavirus aladeformis ), 검은 여왕 세포 바이러스(BQCV, Triatovirus nigereginacellulae ) 및 낭포성 바이러스(SBV, Iflavirus sacbroodi ) 입니다 . 범위 확장의 잠재력이 가장 큰 것으로 제안된 바이러스 스필오버의 주요 예는 A. mellifera 에서 B. impatiens 로의 DWV A 또는 B 마스터 변종의 스필오버입니다 . 야생 Bombus 종은 일반적으로 DWV의 두 마스터 변종 모두에 대해 RT-PCR 양성이며 11 DWV는 적어도 인공 실험 시스템에서 이 속의 일부 구성원에서 복제 주기를 완료할 수 있습니다 12 . 최소한 막다른 스필오버와 DWV 복제가 야생 Bombus 종 에서 감지될 수 있음을 나타냅니다 13 , 14 . 현재 Bombus 에서 지속적인 노출 없이 DWV가 유지된다는 증거는 없습니다 . DWV 및 기타 바이러스에 대한 주요 가설된 스필오버 경로는 공유 꽃에 침전된 바이러스의 경구 섭취를 통한 것이며, A. mellifera 가 방문한 꽃에서 DWV의 RT-PCR 검출을 통해 뒷받침됩니다. 그러나 범위 확장으로 인한 DWV 스필오버에 반대하는 경우도 있습니다. 첫째, 야생 수분매개자에서 제안된 바이러스 스필오버 가 범위 확장으로 이어지는 부정적인 결과에 대한 증거는 거의 없습니다. 10 , 15 다른 연구에서는 DWV 전파가 Bombus 종을 기생하지 않는 Varroa 응애 의 풍부함과 유행과 높은 상관관계가 있고 경구-분변 경로를 통한 DWV 감염은 매우 비효율적이기 때문에 자연적 환경 에서 땅벌 사이 의 성공적인 DWV 전파는 있을 수 없다고 제안합니다. 12 , 16 . DWV 10 에 비해 입수 가능한 증거가 적지만, 다양한 다른 꿀벌 바이러스에 대해서도 스필오버 현상이 제안되었습니다 .
다른 동물 바이러스 스필오버 시스템 7 , 8 의 통찰력을 고려할 때, A. mellifera 에서 다른 곤충으로의 바이러스 스필오버에 대한 증거는 매우 불확실한 것으로 보입니다. 유전체적 접근 방식은 이러한 불확실성 중 일부를 해결할 수 있으며, 특히 새로운 숙주에 적응된 바이러스 변종이 스필오버 이벤트 이후에 출현했는지 여부입니다. 전체 바이러스 커뮤니티의 유전체를 비교함으로써 새로운 숙주 내에서 출현한 변종을 식별할 수 있으며, 이는 다른 곤충에서 A. mellifera 바이러스의 존재가 노출, 막다른 스필오버 또는 범위 확장을 통한 스필오버의 결과인지를 나타낼 수 있습니다. A. mellifera 와 B. impatiens 사이의 바이러스 스필오버 상태를 확인하기 위해 미국 미네소타의 현장 연구 프로그램에서 동소성 A. mellifera 와 B. impatiens 의 메타 전사체를 조사했습니다 . RNA 편향 접근법을 사용하여 우리는 3년 기간에 걸쳐 A. mellifera 바이러스의 B. impatiens 특이적 변종을 식별하여 두 벌에서 확인된 모든 RNA 바이러스에 대한 스필오버 증거를 테스트했습니다 .
결과 및 논의
총 408개의 Apis mellifera 와 131개의 Bombus impatiens 샘플에 대해 시퀀싱 라이브러리를 생성했습니다. A. mellifera 와 B. impatiens 의 경우 각각 19개와 14개의 샘플 라이브러리가 콘티그를 생성하지 않았거나 1000bp 미만의 콘티그만 생성했기 때문에 분석에서 제외되어 총 389개와 117개의 샘플에 시퀀싱 데이터가 있었습니다. A. mellifera 와 B. impatiens 의 경우 라이브러리당 평균 리드 수(Q-점수 임계값을 통과한 리드 수)는 각각 80,563개와 101,004개였고, 평균 리드 길이는 550bp와 487bp였으며, 바이러스 콘티그에 매핑된 리드의 백분율은 각각 9.7%와 4.6%였습니다.
모든 샘플에서 A. mellifera 와 B. impatiens 간의 바이러스 군집은 뚜렷하게 달랐습니다 (PERMANOVA Df = 1, R 2 = 0.007, p = 0.001 ***). A. mellifera 와 B. impatiens 간의 군집 구성 유사성에 유의미한 영향을 미친 유일한 실험 변수는 채집 연도였습니다(PERMANOVA Df = 2, R 2 = 0.01, p = 0.001 ***). 이는 숙주 속과 유의미한 상호 작용을 보인 유일한 변수였기 때문입니다(보충 표 1 ; 보충 그림 1 ).
B. impatiens 의 바이러스 군집은 전반적으로 A. mellifera 보다 샘플링 연도 간에 더 동질적이었습니다 . 두 벌 종을 구별하는 주요 바이러스는 다음과 같습니다(PCA, 그림 2 및 피어슨 상관 계수(>0.1), 보충 그림 2 및 군집 구성, 그림 3으로 평가 ) .A. mellifera : 에서 Cripavirus mortiferum (CM), Lake Sinai 바이러스 3(LSV3), Lake Sinai 바이러스 6(LSV6), 미분류 sinai바이러스(US), Lake Sinai 바이러스 2 (LSV2), Aparavirus apisacutum (AA), Iflavirus sacbroodi (IS). B. impatiens는 분류되지 않은 Vespa Velutina 관련 Permutotetra 유사 바이러스 1(VVPV2), Hymenopteran Phasma 관련 바이러스 OKIAV231(HPVO231), Hubei Picorna 유사 바이러스 27(HPV27), Cyclosorus Interruptus Picorna 유사 바이러스(CIPV) 및 Ganda Orthophasma 바이러스(GO)와 관련이 있습니다. 파스마바이러스과(UP), 메이필드 바이러스 1(MV1), 벌목 파스마 관련 바이러스 OKIAV234(HPVO234), 엘프 로크 바이러스(ELV), 봄부스 관련 바이러스 reo1(BVR1), 아가시즈 록 바이러스(ARV), 박트로세라 트리오니 이플라바이러스 1(BTI1), 봄부스 관련 바이러스 Vir4(BVV4), 안드레나 치질 네게 유사 바이러스(AHNV) 및 Allermuir Hill 바이러스 1(AHV1). 나머지 바이러스는 두 숙주 속(보충 그림 2 )과 중립적 상관관계( r = 0.0 또는 -0.0)를 보였으며 상대적 풍부도가 낮았습니다(그림
관련링크
댓글목록
등록된 댓글이 없습니다.
